nVue-nVision - Technology overview
nVue and nVision together form a high-resolution, multimodal imaging ecosystem that enables the real-time visualization and quantification of neural dynamics and behavior in freely moving animal models. Designed for preclinical neuroscience research, these platforms operate synergistically to provide a comprehensive, temporally precise view of brain activity and behavioral outputs, with millisecond-level resolution and biological relevance across longitudinal studies.
The nVue system is a miniaturized, dual-color fluorescence microscope (miniscope) capable of simultaneously recording optical signals from up to two spectrally distinct genetically encoded indicators. This allows researchers to monitor neuronal activity via calcium imaging using genetically engineered calcium indicators (GCaMP, RCaMP…), blood flow dynamics, or neurochemical fluctuations using genetically engineered neurotransmitter indicators (dopamine via dLight or GRAB sensors) in targeted brain regions. By implanting a GRIN lens into the brain and delivering controlled excitation light through the miniscope, nVue captures fluorescence signals in real time, enabling minimally invasive, longitudinal tracking of circuit-level neural processes in vivo. It supports both static imaging of a specific cell population in a defined brain areas and dynamic measurements of localized physiological activity.
The nVision platform complements nVue by integrating its neural imaging output with high-definition behavioral tracking, real-time experimental control, and environmental modulation. Using DeepLabCut’s AI-enhanced video analysis, nVision enables synchronized capture of locomotion, posture, social interactions, and task-specific behaviors, correlating them precisely with concurrent neural activity. The platform also supports closed-loop paradigms, where defined neural events can automatically trigger external stimuli (light, sound, or reward delivery), allowing for causal investigations into brain-behavior relationships.
Together, nVue and nVision empower researchers to perform longitudinal, systems-level investigations, bridging the gap between molecular signaling, neural circuit function, and complex behaviors. These tools are particularly advantageous in fields such as cognitive neuroscience, behavioral pharmacology, and neurological disease modeling, where understanding the dynamic interplay between neural activity and behavior is critical.
Important considerations
Miniature fluorescence microscopy enables high-resolution, real-time imaging of neural activity in freely behaving rodents. Its effectiveness critically depends on the use of genetically encoded biosensors, specifically genetically encoded calcium indicators (GECIs) and genetically encoded neurotransmitter indicators (GENIs). These molecular tools transduce intracellular calcium transients and extracellular neurotransmitter fluctuations, respectively, into fluorescent signals. When expressed in targeted neuronal populations, they permit cell-type-specific, spatiotemporally precise visualization of circuit dynamics underlying behavior, learning, and neuromodulation. For additional details regarding fluorescence, please refer to the section Fluorescence imaging.
Intracellular calcium biosensors
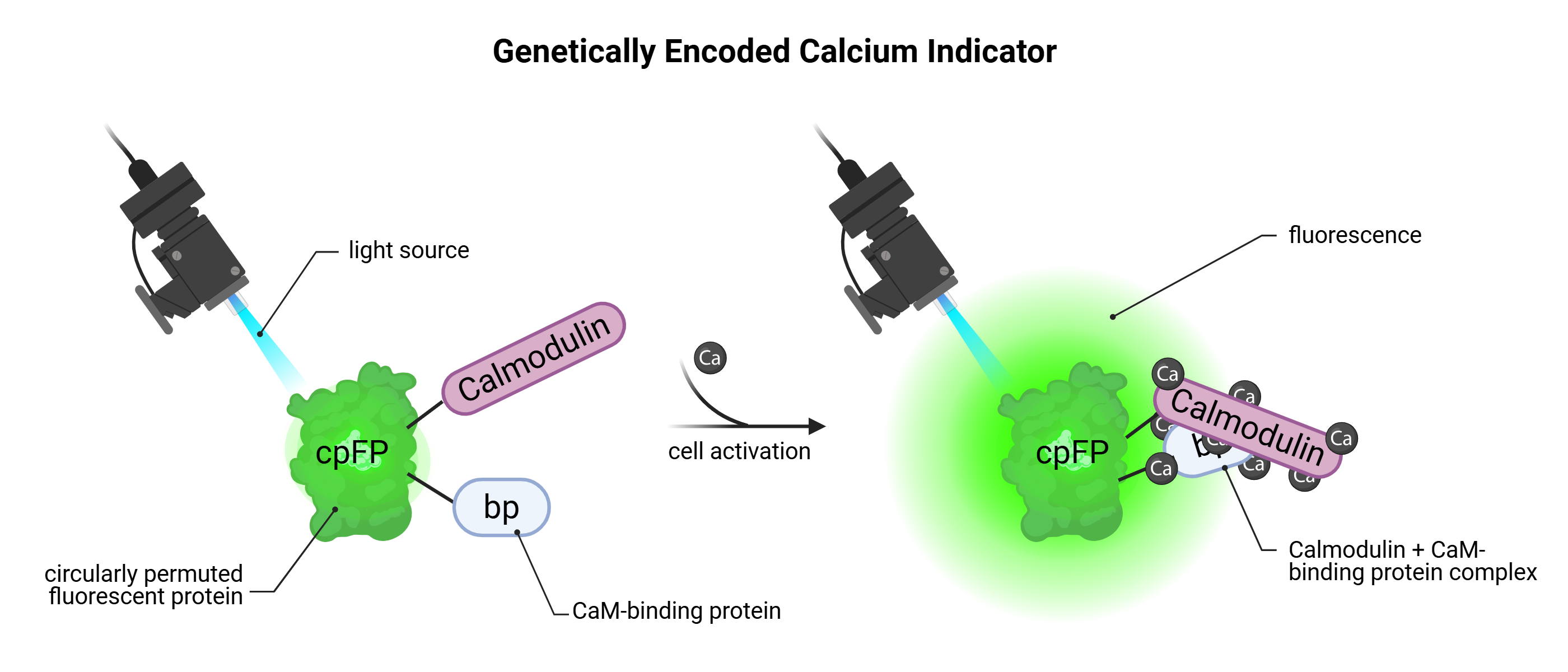
Mechanism of Genetically Encoded Calcium Indicators (GECIs)
Genetically encoded calcium indicators (GECIs) are cytoplasmic molecular biosensors engineered to optically report changes in intracellular calcium concentrations ([Ca²⁺]ᵢ), which serve as reliable and indirect proxies for neuronal activity. The most extensively used class of GECIs is the GCaMP family, which consists of a circularly permuted green fluorescent protein (cpGFP) fused to calmodulin (CaM) and a calmodulin-binding peptide (M13). During neuronal depolarization, the opening of voltage-gated calcium channels permits calcium influx. Binding of calcium to calmodulin promotes a conformational rearrangement that facilitates its interaction with M13, ultimately stabilizing the cpGFP chromophore and resulting in a pronounced increase in fluorescence intensity. This calcium-dependent fluorescence response is both rapid and reversible, enabling the real-time monitoring of neuronal dynamics with high temporal fidelity. Due to their protein structure, GECIs exhibit baseline fluorescence even in the absence of calcium binding, stemming from the constitutive folding of cpGFP. While this background signal is advantageous for confirming expression and maintaining optical traceability, it imposes a limitation on the signal-to-noise ratio, particularly in circuits with low spiking frequency or subthreshold activity. Consequently, accurate interpretation of fluorescence traces necessitates rigorous calibration, thresholding, and often the selection of high-dynamic-range variants, such as GCaMP6f or GCaMP7f, which are specifically optimized for detecting fast, transient signals associated with single or few action potentials.
A diverse portfolio of GCaMP variants has been developed, each offering distinct kinetic profiles and sensitivity ranges, thereby enabling researchers to match the indicator to the demands of the experimental paradigm, from millisecond-resolved detection of individual spikes to integrated monitoring of population-level oscillations. When expressed under cell-type-specific or region-specific promoters, or delivered via viral vectors, GECIs allow for targeted, genetically precise imaging of neural ensembles.
Altogether, the expanding library of GECI tools, ranging from ultra-fast kinetic sensors to integrative, high-sensitivity variants, offers unparalleled flexibility for studying brain function. Their use has become foundational in modern systems neuroscience, particularly for interrogating the spatiotemporal organization of neural circuits during behavior, learning, memory, and disease states.
Synaptic neurotransmitter biosensors
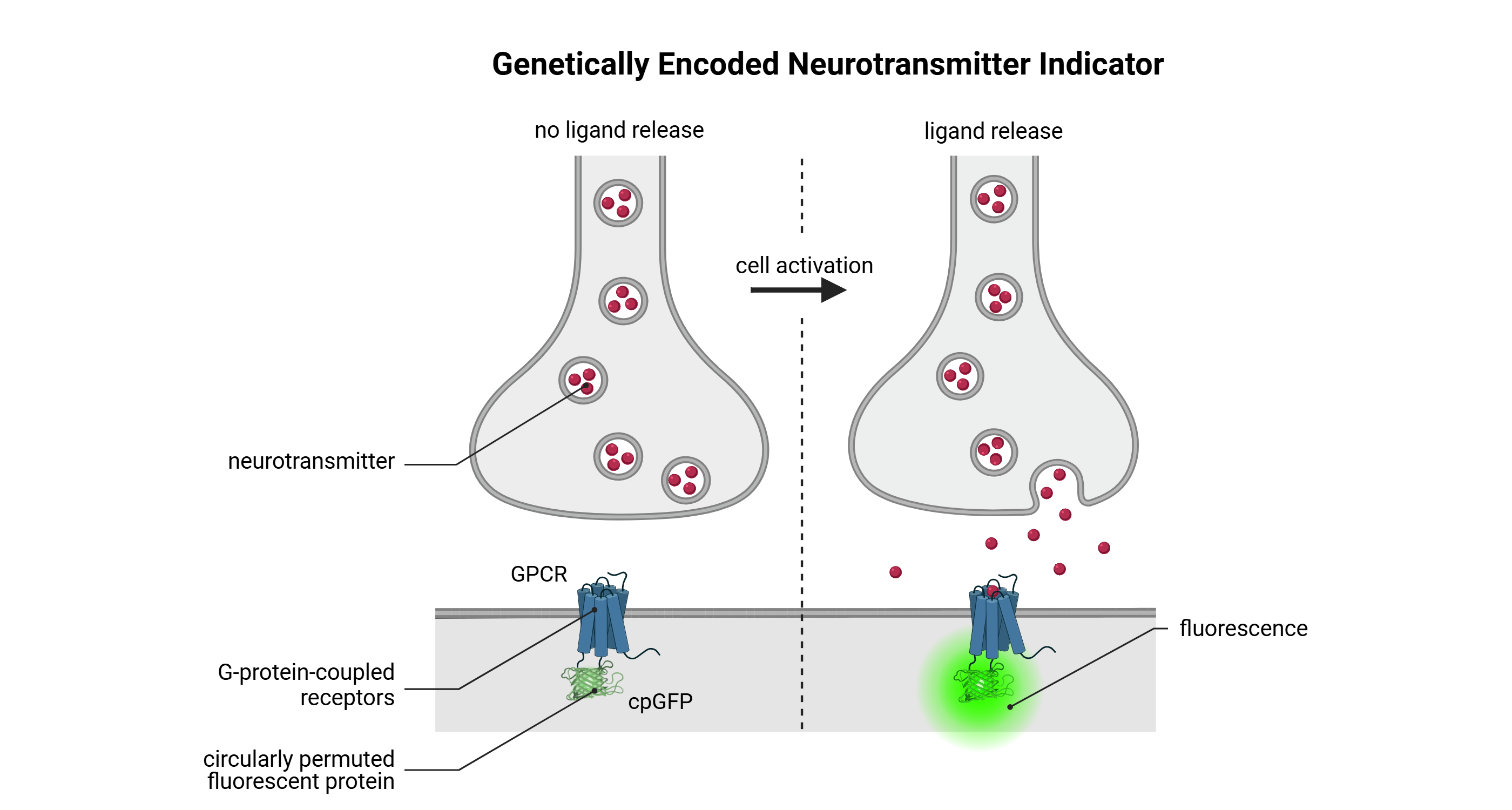
Mechanism of action of Genetically Encoded Neurotransmitter Indicator (GENIs)
Genetically encoded neurotransmitter indicators (GENIs) are membrane-localized molecular biosensors engineered to optically report fluctuations in extracellular neurotransmitter concentrations, thereby enabling the real-time detection of synaptic transmission and neuromodulatory events in vivo. These sensors are typically constructed by fusing a neurotransmitter-binding domain, often derived from modified G protein-coupled receptors (GPCRs), to a circularly permuted fluorescent protein (commonly cpGFP). Upon ligand binding, the GPCR-derived domain undergoes a ligand-induced conformational rearrangement, which alters the environment of the cpGFP chromophore and leads to a robust, reversible change in fluorescence intensity. This molecular mechanism enables the transformation of transient chemical signals into quantifiable optical readouts with high spatial and temporal precision.
GENIs are available for a broad range of neurotransmitters, including dopamine (dLight1, GRAB-DA), glutamate (iGluSnFR), acetylcholine (GRAB-ACh), serotonin (GRAB-5HT), and others. Their performance is defined by tunable parameters such as ligand affinity, dynamic range, response kinetics, and photostability, which can be optimized to match specific physiological release patterns, including phasic versus tonic signaling. These indicators are particularly well-suited for monitoring neuromodulatory dynamics in targeted brain regions, offering critical insights into the cellular and circuit-level mechanisms underlying reward processing, motivation, arousal, learning, and behavioral flexibility.
When delivered via viral vectors and expressed under cell-type- or projection-specific promoters, GENIs allow for precise, circuit-defined measurement of neurotransmitter signaling in both local and long-range pathways. Similar to GECIs, GENIs exhibit a degree of baseline fluorescence in the absence of ligand, attributable to the structural conformation of the cpGFP domain. While this background signal facilitates monitoring of expression levels and cellular localization, it necessitates appropriate calibration and normalization to distinguish authentic neurotransmitter-evoked responses from baseline fluctuations. High-sensitivity variants, such as dLight1.3b and GRAB-DA2m, have been engineered to optimize detection across physiological concentration ranges, allowing for subsecond resolution of neuromodulatory events during naturalistic behavior.
Comparison of GECI and GENI

Comparison of GECIs and GENIs biosensors
Miniscope imaging
Hardware
A miniscope is a miniaturized epifluorescence microscope engineered to be compact and lightweight, allowing it to be mounted onto the head of a freely moving mouse with minimal discomfort or disruption of natural behavior.
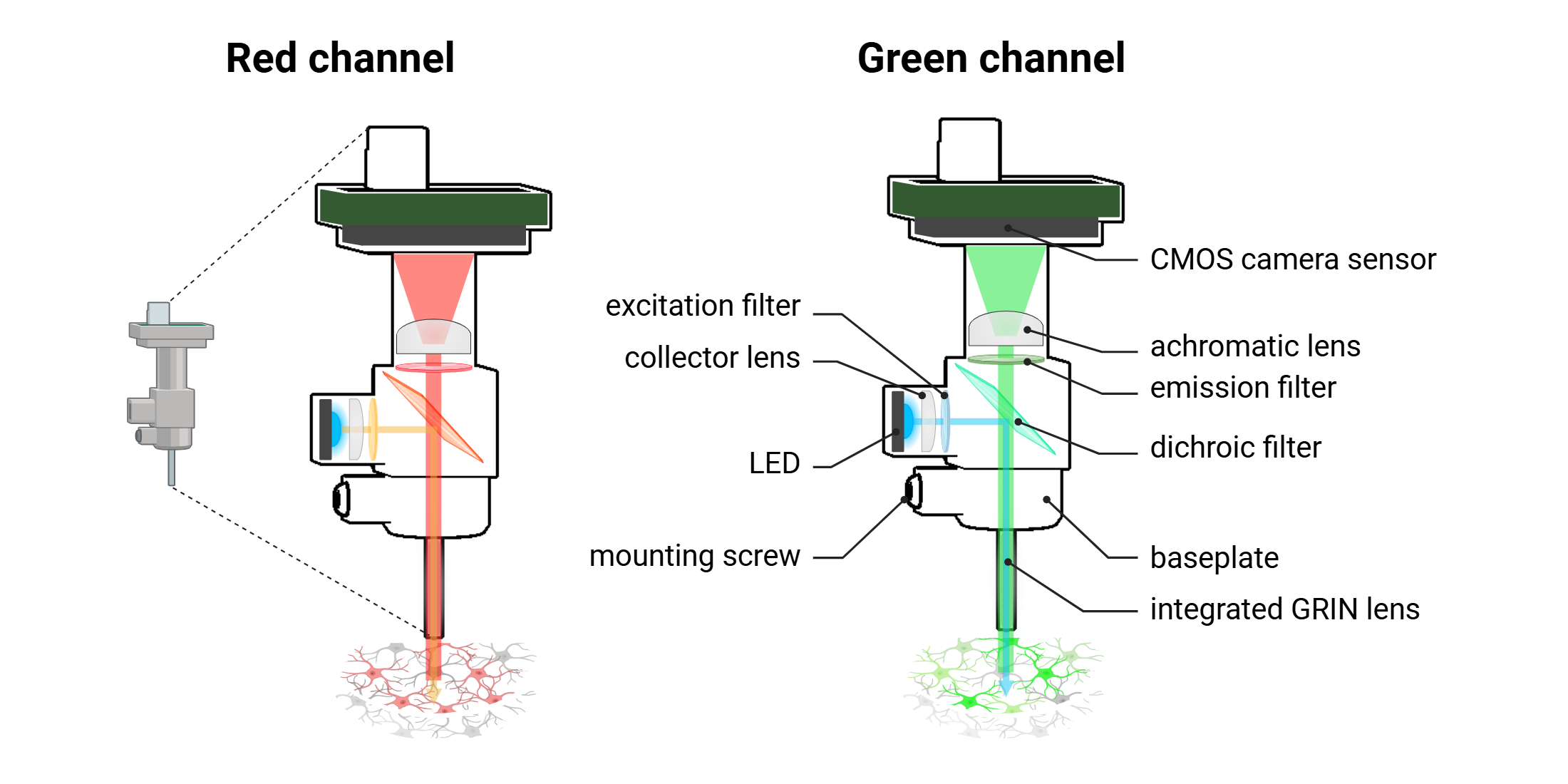
In vivo dual color miniscope imaging
As with conventional fluorescence microscopes, the miniscope includes an excitation light source, (high-power LED). The divergent light emitted from the LED is first directed by a collector lens, which focuses and channels the beam into the optical system. This incident light is then passed through an excitation filter, which spectrally selects the appropriate wavelength required to excite the targeted fluorescent reporter. The refined excitation light is reflected downward by a dichroic mirror, which is specifically designed to reflect excitation wavelengths while transmitting longer-wavelength emitted fluorescence. The directed excitation light then enters the brain through the objective lens, which in miniscopes is typically a gradient-index (GRIN) lens. This GRIN lens has a radially graded refractive index, allowing it to efficiently focus the excitation light into deep brain structures while maintaining compact form and minimal optical aberration. Upon excitation, fluorophores expressed in neurons or surrounding tissue emit photons corresponding to their characteristic emission spectra. These emitted photons travel back through the same GRIN lens and are transmitted through the dichroic mirror, which now functions as a pass-through filter for the emission wavelengths. The fluorescence signal is subsequently passed through an emission filter, which further isolates the desired emission band while eliminating residual excitation light. Finally, the filtered emission light is directed through an achromatic imaging lens, which focuses the signal onto a CMOS camera sensor positioned at the distal end of the miniscope. This configuration enables the acquisition of a high-resolution, two-dimensional fluorescence image that reflects the spatial distribution and temporal dynamics of neural activity within the defined field of view. The achromatic lens corrects for chromatic aberration across the emission spectrum, ensuring sharp image quality and signal fidelity across different fluorescence channels.
GRIN lens
Gradient Index (GRIN) lenses are optical elements that exploit a radial gradient in refractive index to bend light rays internally rather than at curved surfaces, as in traditional lenses. As a result, GRIN lenses typically feature flat entry and exit faces and are designed to guide light along a curved path within the lens body. In miniscope imaging systems, GRIN lenses function as 1:1 relay lenses, meaning they transmit the image plane along their length without altering magnification. Their primary purpose is to extend the optical path into deep brain regions while preserving resolution and contrast.

GRIN lens geometry
There are two main types of GRIN lenses used in neural imaging, and selection should be based on the target brain structure and orientation of the neuronal population:
Straight GRIN lenses: These lenses transmit the image directly along the longitudinal axis of the probe. They are ideal for imaging neuronal layers positioned directly beneath the implanted lens, and are the most commonly used type in in vivo experiments targeting structures such as the hippocampus, thalamus, striatum, or hypothalamus.
Prism GRIN lenses: These incorporate a 90-degree reflective prism at the distal tip, enabling lateral imaging perpendicular to the insertion axis. This design allows for visualization of structures that are located adjacent to the probe body, rather than directly below it. Prism lenses are particularly advantageous for cortical imaging, as they permit multi-layer field-of-view acquisition without needing to penetrate vertically through all cortical layers.
Because GRIN lenses are made of precision optical glass, their imaging surfaces must be handled with care. Any scratches, chips, adhesive residue, or surface contamination can degrade optical performance, introduce aberrations,or obstruct the field of view. If needed, clean the lens using lens paper moistened with 70% ethanol, applying gentle, circular motions to avoid damaging the surface. Avoid touching the imaging face with bare tools or gloves, and always cover the lens when not in use to prevent environmental contamination.
Workflow for in vivo miniscope imaging
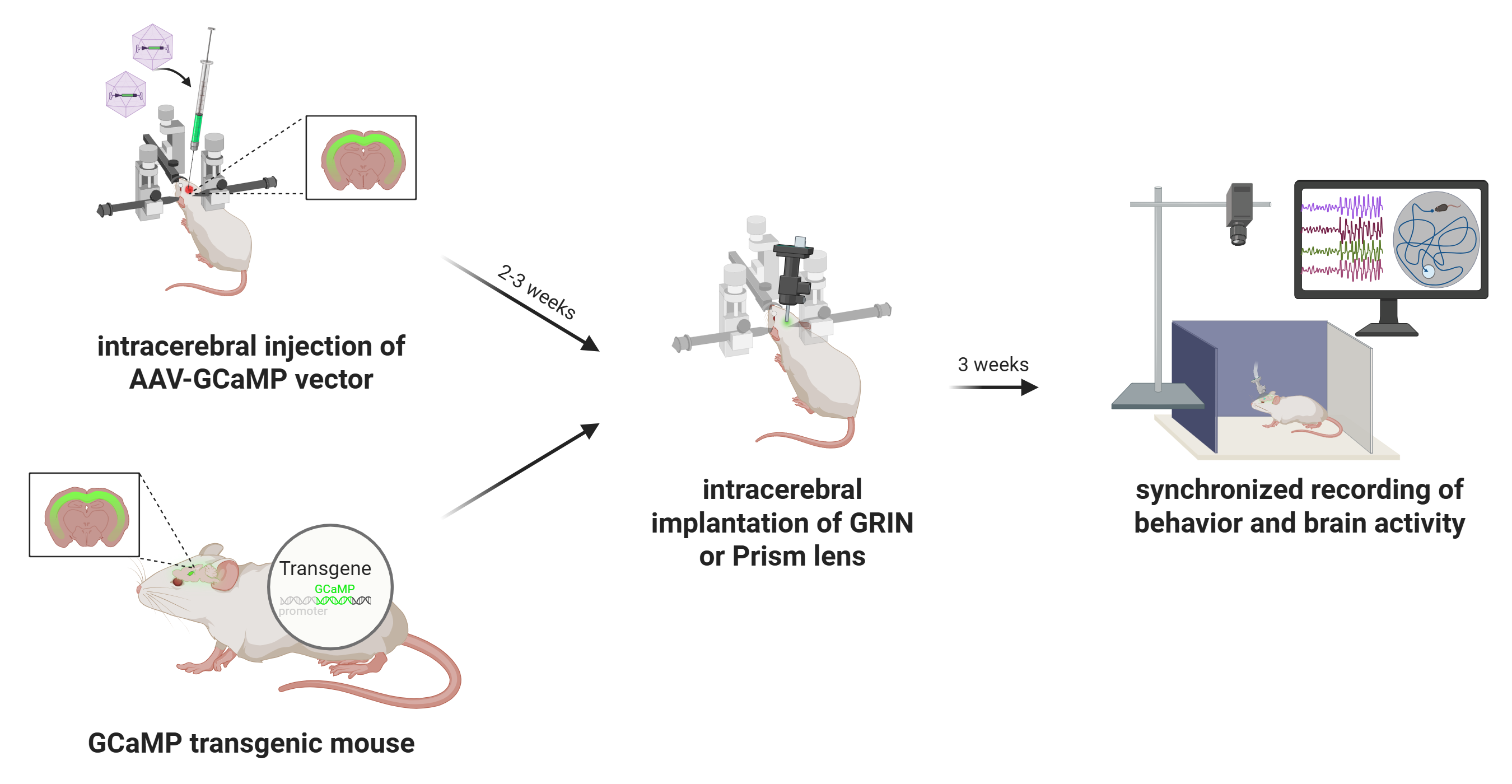
Common workflow for in vivo miniscope imaging with dual color
The successful implementation of miniscope-based imaging in freely behaving animals requires a multi-stage experimental workflow that integrates genetic targeting of fluorescent indicators, surgical implantation of a gradient-index (GRIN) lens, and chronic in vivo optical recording. Each phase must be carefully optimized to ensure high-quality data acquisition and minimal disruption to animal health or behavior.
Targeted expression of fluorescent biosensors
Miniscope imaging relies on the expression of genetically encoded calcium indicators or neurotransmitter indicators in neurons of interest. This is typically achieved through one of two strategies:
AAV-mediated viral delivery
Recombinant adeno-associated virus (AAV) vectors are commonly employed to drive the expression of genetically encoded fluorescent indicators in targeted neuronal populations. The indicator gene is placed under the control of a cell-type-specific or pan-neuronal promoter, such as human synapsin (hSyn) for broad neuronal expression, or CaMKIIα for excitatory neuron selectivity. AAV constructs are delivered via stereotaxic injection into the brain region of interest (hippocampus, striatum, or cortex), with precise spatial targeting guided by anatomical landmarks or coordinates derived from a stereotaxic atlas. The choice of AAV serotype (AAV1, AAV5, AAV9) and viral titer must be carefully optimized to achieve efficient cellular transduction while minimizing cytotoxic effects, inflammatory responses, and off-target expression. Optimization of viral titer, serotype, and targeting parameters is typically validated through direct visualization of the fluorescent reporter in brain tissue. In many cases, when the indicator is expressed at sufficient levels, native fluorescence can be readily observed using standard fluorescence microscopy in either fresh brain slices or fixed tissue. This approach allows rapid, non-destructive assessment of expression strength, anatomical accuracy, and spread of transduction, and is especially useful for high-throughput evaluation across experimental animals. In addition to native fluorescence imaging, post-mortem immunohistochemistry may be employed for enhanced sensitivity or when expression levels are low. IHC uses antibodies targeting the reporter protein (anti-GFP for GCaMPs) or a co-expressed fluorescent tag (mCherry, tdTomato), allowing high-resolution evaluation of cell-type specificity, transduction efficiency, and signal localization. Together, these validation approaches ensure that the chosen AAV construct and delivery parameters result in optimal, stable biosensor expression prior to GRIN lens implantation and chronic imaging.
Following AAV injection, a 2–4 week expression window is generally required to allow sufficient indicator production and functional folding. This period also ensures stabilization of fluorescence intensity and minimizes confounding effects due to acute viral response or surgical trauma. Only after this maturation window is it advisable to proceed with GRIN lens implantation and functional imaging using the miniscope platform.
Transgenic animal models
Transgenic mouse lines expressing genetically encoded calcium indicators (Thy1-GCaMP6, TIT2L-GCaMP6f) offer a robust and reproducible alternative to viral vector-based expression systems. These lines provide stable, heritable, and cell-type-specific expression of calcium indicators under the control of defined promoters, enabling consistent signal intensity and anatomical coverage across animals and experiments. Importantly, the use of transgenic models eliminates the need for stereotaxic AAV injection, thereby reducing surgical burden and avoiding potential sources of experimental variability. Transgenic GCaMP lines are particularly well-suited for long-term, chronic imaging studies, developmental time-course experiments, and investigations requiring minimal invasiveness or inter-animal variability. These advantages are especially valuable in behavioral neuroscience, where consistent expression across cohorts is essential for meaningful comparisons. However, their use is inherently constrained by the available genetic constructs, limiting flexibility in targeting specific cell types, projection-defined populations, or combinatorial expression strategies. As such, while transgenic lines provide a valuable platform for standardization and scalability, their application must be weighed against the need for targeted circuit-level specificity, which is more readily achieved through AAV-mediated delivery in combination with Cre/Flp recombinase systems.
Intracranial implantation of the GRIN lens
Following the establishment of robust biosensor expression, a baseplate-integrated GRIN lens is surgically implanted under stereotaxic guidance to provide chronic optical access to the brain region of interest. The procedure begins with a craniotomy over the targeted area, using stereotaxic coordinates for precise anatomical targeting. The GRIN lens is then carefully positioned approximately 150 μm above the fluorescently labeled neuronal population, thereby avoiding mechanical disruption of active neural tissue while preserving optimal optical coupling. Once in place, the lens and integrated baseplate are permanently secured to the skull, forming a stable interface for subsequent attachment of the head-mounted miniscope. Importantly, optimization of GRIN lens implantation requires post-mortem histological analysis to verify implantation coordinates, confirm targeting accuracy, and assess potential tissue disruption. This step is essential for validating anatomical precision and for interpreting imaging data in the context of localized neural circuits.
Following surgery, animals are allowed to recover for 1–2 weeks to support wound healing, tissue stabilization, and attenuation of glial reactivity. This recovery period also allows the optical interface between the lens and brain tissue to equilibrate, thereby enhancing imaging clarity and ensuring consistency for longitudinal, high-resolution recordings in freely behaving animals.
Miniscope mounting and imaging
Once the GRIN lens is stably integrated and fluorescent reporter expression has been verified, the head-mounted miniscope is secured to the implanted baseplate to initiate in vivo imaging. Prior to each imaging session, key acquisition parameters (LED excitation power, sensor gain, and electronic focal depth…) are adjusted using dedicated acquisition software to optimize the signal-to-noise ratio while minimizing photobleaching and phototoxicity. Imaging is performed while the animal is awake and freely behaving, either during spontaneous exploration or in structured behavioral paradigms including open field tests, maze-based navigation, or operant conditioning tasks. Animal behavior is simultaneously recorded using synchronized video tracking systems, enabling precise temporal alignment between neural activity and behavioral events. Fluorescence signals are acquired at frame rates typically ranging from 20 to 60 Hz for calcium imaging, depending on the experimental demands and the kinetic properties of the biosensor used. Both single-channel and dual-channel acquisition modes are supported, enabling simultaneous measurement of multiple biological signals, such as neuronal calcium dynamics and neurotransmitter release, within the same field of view. Imaging sessions can be repeated longitudinally over days to weeks, allowing for the chronic tracking of identified neuronal populations, investigation of circuit-level plasticity, and mapping of behaviorally relevant neural activity patterns across a wide range of experimental conditions.
Data analysis
Following data acquisition, raw miniscope imaging videos undergo a structured preprocessing and analysis pipeline to extract biologically meaningful neural activity traces. The first step involves motion correction, which compensates for brain movement and animal locomotion using frame-by-frame image registration algorithms that align frames to a common reference. This step is essential for maintaining spatial fidelity of fluorescence signals over time. After motion correction, the videos are subjected to spatial cropping and temporal downsampling, depending on the experimental needs, followed by background subtraction to enhance signal contrast. Next, ROIs (neuronal soma) are identified using semi-automated or fully automated cell segmentation algorithms, including approaches based on PCA/ICA, CNMF-E (constrained non-negative matrix factorization), or machine learning-based classifiers. These segmented ROIs are then used to extract raw fluorescence traces (F), which are typically normalized to obtain ΔF/F₀ values, calculated as ΔF/F₀ = (F - F₀) / F₀, where F₀ represents the baseline fluorescence. This normalization facilitates comparison across neurons and imaging sessions. Additional processing steps include deconvolution to estimate underlying spike trains, trace denoising, and quality control filtering to exclude low-SNR signals or artifactual ROIs. Once individual cell traces are obtained, downstream analyses may include population dynamics, activity correlations, event-triggered averages, and behavioral alignment based on timestamp synchronization with external behavioral data (locomotion, trial events, video-tracked metrics). These analyses provide critical insight into how neural activity encodes stimuli, behavior, learning, and circuit-level computations over time.
Advantages of miniscope imaging
High spatial resolution at the cellular level
Miniscopes enable single-neuron resolution through head-mounted optics and GRIN lens technology. These systems typically achieve lateral spatial resolution in the range of 1–2 μm, which is sufficient to resolve individual somata and, in some cases, dendritic segments. This cellular resolution is critical for identifying and tracking discrete neuronal units across time, enabling longitudinal studies of plasticity, learning, or disease progression. Combined with proper lens alignment and stable implantation, the system provides a stable optical interface for high-fidelity signal capture within deep brain regions.
Dual-color fluorescence imaging for enhanced specificity
nVue supports simultaneous imaging of two fluorescent channels, enabling precise discrimination of cell types or activity signals. Dual-color imaging improves the biological specificity of the data by allowing concurrent visualization of distinct cell populations or genetically encoded sensors. This multiplexing capacity enhances interpretability in circuit-level experiments, such as distinguishing inhibitory versus excitatory neuron dynamics or separating calcium and neurotransmitter activity. Dual-color capabilities also reduce the need for cross-animal comparisons, improving within-subject control and reducing inter-sample variability.
**High sensitivity for detecting calcium transients
The system achieves excellent sensitivity by combining high-power, wavelength-matched LED excitation with low-noise CMOS sensors. It is capable of capturing small changes in intracellular calcium levels, which reflect neuronal spiking activity. When paired with bright, genetically encoded indicators, the system offers reliable detection of subthreshold and suprathreshold activity, even in low-expressing neurons. This sensitivity is critical for decoding sparse or weakly modulated signals in behaviorally relevant contexts.
Robust SNR under freely behaving conditions
Imaging during unrestrained behavior introduces motion and optical variability, but the miniscope design minimizes these effects. The use of fixed focal GRIN lenses, optimized optics, and active motion correction (during post-processing) supports a high signal-to-noise ratio, even under conditions of moderate locomotion. Frame rates of up to 40 fps help temporally isolate neural events from movement artifacts, and spatial filtering algorithms further enhance SNR. This allows for the extraction of clean fluorescence traces critical for spike inference and population decoding.
Longitudinal tracking of neural ensembles
Miniscope implants allow for stable imaging across days to weeks from the same field of view. The baseplate system ensures consistent alignment of the optical axis across imaging sessions, enabling precise re-identification of neuronal ROIs over time. This longitudinal capability supports studies of experience-dependent plasticity, learning trajectories, neurodegeneration, and recovery. Importantly, signal integrity can be maintained across sessions with careful baseplate handling, ensuring that changes in neural activity reflect true biological phenomena rather than sampling error or alignment drift.
Integration with behavioral and electrophysiological systems
nVision software enables tight synchronization between imaging and behavior or stimulation paradigms. The miniscope system includes TTL input/output and timestamped metadata logging, allowing integration with video tracking, behavioral software, or concurrent electrophysiology. This allows researchers to align neuronal dynamics with precise behavioral events, such as reward delivery, stimulus onset, or motor transitions. Such synchronization is essential for decoding context-dependent neural activity and performing causality-based analyses (peri-event histograms or population trajectory modeling).
Lightweight design for naturalistic behavior
Miniscopes are designed to preserve normal locomotion and exploratory behavior. Weighing under 3 grams, they minimizes head load and mechanical interference, making it suitable for mice and juvenile rats. The lightweight housing ensures that locomotion, rearing, grooming, and other behaviors remain natural and unaffected, reducing confounds due to handling or restraint. This feature is particularly important in behavioral neuroscience, where ecological validity is essential for accurate interpretation.
Limitations of miniscope imaging
Restricted field of view limits spatial coverage
Miniscope systems are constrained to small imaging windows, typically around 600–800 μm per channel. This limited FOV restricts the number of neurons that can be imaged simultaneously to a few hundred, depending on cell density and expression level. As a result, the system captures only a partial representation of the local circuit, which may not fully reflect network-level dynamics. This can impact interpretations in studies involving distributed processing, population coding, or inter-regional coordination, where access to broader spatial coverage is essential.
Invasive lens implantation causes tissue disruption
Recording requires surgical implantation of a GRIN lens directly above the imaging target. GRIN lens implantation displaces overlying tissue and may trigger inflammatory responses, gliosis, or damage to local vasculature. These biological consequences can alter baseline neural activity or long-term circuit function. Furthermore, precise targeting is critical: if the lens is misaligned or placed too deep, the imaging plane may not coincide with the desired cellular layer, reducing spatial specificity and effective resolution.
Limited imaging depth and volume
Only a narrow axial band (~100-300 μm below the lens tip) is accessible for functional imaging. Because miniscopes rely on epifluorescence imaging through a static focal plane, optical sectioning is minimal and depth discrimination is limited. This restricts recordings to a single focal layer at a time and precludes volumetric imaging without re-implantation or hardware modification. Thus, miniscope imaging lacks the depth resolution and 3D coverage offered by two-photon microscopy or light-sheet techniques.
Low optical sectioning and axial resolution
Miniscopes rely on widefield epifluorescence without depth discrimination. Unlike two-photon or confocal systems that reject out-of-focus light, miniscopes collect fluorescence from the entire illuminated column. This results in optical blur from neuropil and non-target layers, reducing axial resolution and compromising specificity in densely labeled regions. Signal crosstalk from overlapping cells can lower the accuracy of ROI segmentation and calcium trace interpretation.
Susceptibility to motion artifacts in freely moving animals
Head movement, grooming, or collisions with the environment introduce optical instability, which is particularly problematic when targeting deep brain regions. In freely behaving animals, mechanical forces can cause subtle shifts or rotational jitter of the miniscope, reducing signal fidelity and introducing spatial misalignment across frames. These artifacts lower the effective SNR and impair both single-trial trace extraction and longitudinal cell tracking. The challenge is even more pronounced when imaging deep brain structures where GRIN lenses must traverse large volumes of brain tissue. In such cases, even minor relative motion between the lens and target neurons can disrupt focal alignment, increase signal crosstalk from non-target planes, and reduce spatial resolution.
Risk of photobleaching and phototoxicity
Prolonged or high-intensity fluorescence excitation can damage tissue, degrade signal quality, and alter neural physiology. Miniscope imaging depends on sustained exposure to excitation light, typically via high-intensity LEDs. Over time, this exposure increases the risk of photobleaching, reducing fluorescence signal amplitude and dynamic range, especially in genetically encoded calcium indicators. Additionally, phototoxicity from light-induced heat and oxidative stress may cause tissue inflammation or perturb neural activity. These effects are particularly problematic during long recording sessions or when using higher excitation powers to boost sensitivity. To mitigate these risks, it is strongly recommended to limit continuous imaging to 15 minutes per session, and if longer recordings are needed, to implement a 5-minute recovery break every 15 minutes. These pauses allow for tissue cooling and fluorophore recovery, thereby preserving both SNR and tissue integrity across extended imaging protocols.
High technical demands for surgical preparation
Successful imaging requires precise stereotaxic targeting and advanced surgical technique. Craniotomy, viral injection and GRIN lens implantation must all be executed with sub-millimeter accuracy. Errors at any stage can compromise imaging quality or render animals unusable. Moreover, post-operative care is intensive and critical for reducing inflammation and ensuring lens clarity. As a result, extensive training and pilot testing are needed before deploying the technique at scale.
Requirement for region- and cell-type specific optimization
Effective miniscope imaging demands tailored optimization of injection and implantation procedures based on the targeted brain region and neuronal population. Unlike generic imaging approaches, miniscope-based calcium imaging depends on precise anatomical targeting and robust indicator expression in the region and cell type of interest. Each brain structure has unique stereotaxic coordinates, anatomical constraints, and cellular organization, requiring customized viral injection protocols. Parameters such as anteroposterior, mediolateral, and dorsoventral coordinates, as well as injection angle, vector titer, volume, serotype, and promoter specificity must be carefully optimized for each experiment. Similarly, GRIN lens implantation depth and trajectory must be adjusted to account for the structure’s geometry and the optimal focal offset for fluorescence imaging. Suboptimal targeting can result in weak or off-target expression, insufficient signal-to-noise ratio, or imaging planes misaligned with active neuronal layers. Therefore, preliminary pilot experiments, combined with post hoc validation via histology or fluorescence microscopy, are essential to refine coordinates and ensure reproducibility across animals and cohorts.
Indirect readout of neural activity
Miniscopes detect calcium signals, which are proxies (not direct measures) of spiking. Calcium imaging provides a delayed and integrated signal of action potential activity, with temporal resolution limited by indicator kinetics. While sufficient f or many behaviors, this makes it difficult to resolve rapid spike timing or subthreshold events. Deconvolution algorithms help estimate spike trains but cannot fully replace the temporal precision of electrophysiological recordings.
High data storage and processing requirements
Each session generates large raw datasets requiring robust infrastructure. High-frame-rate recordings across multiple sessions quickly accumulate to hundreds of gigabytes per animal. Efficient data management, compression, and backup strategies are essential. Moreover, image preprocessing (motion correction, segmentation, trace extraction) is computationally demanding and often time-consuming, creating a bottleneck for high-throughput experimental workflows.